К местным анестетикам относятся соединения, вызывающие обратимый блок потенциалов действия нерва. Ряд соединений со свойствами местных анестетиков существуют в природе, включая кокаин и эугенол, добываемые из растений, сакситоксин, получаемый из водорослей (динофлагелляты), и тетродотоксин, извлекаемый из некоторых видов рыб семейства Tetradoniformes (хотя на самом деле это токсин, продуцируемый бактерией Pseudoalteromonas внутри рыб). Нет сомнений в том, что первобытные люди использовали их столетиями, но сообщение о первом применении препарата в качестве местного анестетика относится к 1884 г., когда немецкий врач Карл Коллер применил кокаин, о котором узнал от Зигмунда Фрейда, для топической анестезии в офтальмологии.
В связи с потенциальной токсичностью кокаина химики попытались синтезировать заменитель кокаина еще в начале 1890-х годов. Результатом этих попыток стал синтез прокаина Эйнгорном с соавторами в 1905 г. Все местные анестетики, доступные в настоящее время для регионарной анестезии, являются более эффективными вариантами прокаина.
Химические свойства
- Структура. Все местные анестетики, применяемые для блокад нервов, состоят из гидрофобного ароматического кольца, соединенного с третичной аминогруппой углеродной цепочкой. Длина углеводородной цепочки варьирует от 6 до 9 А; препараты, цепочка которых длиннее или короче, неэффективны. У бензокаина, применявшегося только для топической анестезии, нет третичной аминогруппы, и он не имеет водорода, который заменяется при физиологическом pH (рКa = 3,5).
- Эфиры и амиды. Местные анестетики делятся на эфиры и амиды в зависимости от того, присоединена ли углеродная цепочка к бензольной части эфирной или амидной связью. Тип связи важен для определения того, по какому пути пойдет метаболизм препарата.
- Изомеры. Многие местные анестетики имеют по крайней мере один асимметричный атом углерода и, таким образом, существуют в виде двух или более энантомеров. Большинство из них применяются клинически в виде рацемических смесей, содержащих оба энантомера. Исключение составляют ропивакаин и левобупивакаин: они поставляются в виде одного энантомера, поскольку в клинической практике он действует сильнее и менее токсичен, чем рацемат.
Физико-химические свойства
А. Кислота-основание. Поскольку третичные аминогруппы могут связывать протон, чтобы стать позитивно заряженным четвертичным амином, все местные анестетики (за исключением бензокаина) существуют в виде слабой пары кислота-основание в растворе. Способность генерировать положительный заряд критически важна для блокады натриевых каналов.
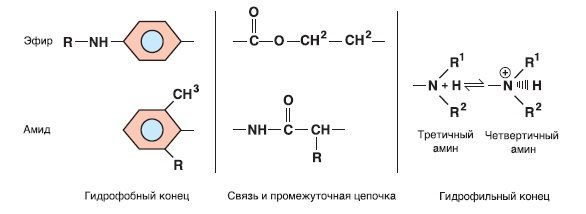
Типичная структура молекул местных анестетиков эфиров и амидов
1. Константа диссоциации рКa. В растворе местные анестетики существуют как в незаряженной форме (основание), так и в положительно заряженной (кислота). Доля каждого из них в конкретном растворе или ткани зависит от pH раствора/ткани и его можно рассчитать по уравнения Гендерсона—Хассельбаха (Henderson—Hasselbalch):
рКa = pH — log [основание]/[кислота],
где pH — pH раствора или ткани и рКa — pH при которой половина молекул местного анестетика присутствует в форме основания, и половина — в форме кислоты.
Значение рКa уникально для каждого местного анестетика и определяет тенденцию молекулы присоединять протон, когда она находится в форме основания, или отдавать протон, когда она в форме кислоты. Большинство местных анестетиков имеют рКa в диапазоне от 7,5 до 9,0.
Поскольку местные анестетики поставляются в несбуфферированном кислом растворе (pH = 3,5—5,0), в них от 1000 до 100 000 раз больше молекул в заряженной в форме, чем в незаряженной неионизированной (что способствует сохранности местного анестетика в растворе). Поскольку pH внеклеточных тканей равен приблизительно 7,4, при инъекции в ткани пропорция заряженных молекул под действием этого фактора снижается в 500—10 000 раз. Например, так как рКa мепивакаина равна 7,6, в коммерчески доступном растворе с pH 4,6 должно быть в 1000 раз больше молекул в протонной форме (слабая кислота), чем в незаряженной. После инъекции в ткани с pH 7,4 многие заряженные молекулы мепивакаина должны «отдавать» свои протоны, и теперь заряженных будет больше, чем незаряженных, приблизительно в 1,6 раза. Способность переходить из заряженной формы в незаряженную и обратно критически важна для действия местного анестетика.
Физико-химические свойства местных анестетиков
Способность местных анестетиков растворяться в водных (гидрофильных) и жировых (гидрофобных) средах варьирует. Различия по гидрофобности возникают в первую очередь как следствие различий типов химических групп связывания с четвертичными аминами. Заряженная форма любого местного анестетика более гидрофильна, чем соответствующая незаряженная форма. Гидрофобный характер часто обозначают как жирорастворимость — что не точно. Более выраженная гидрофобность коррелирует с более выраженной мощностью местного анестетика и длительностью его действия.
1. Гидрофобность определяется добавлением местного анестетика в сосуд, содержащий две несмешиваемых жидкости — водного буфера и гидрофобного «липида». Липиды обычно выбирают так, чтобы замаскировать гидрофобный характер липидных мембран клеток; часто используют октанол, оливковое масло и п-гептан. Местный анестетик добавляют в сосуд и встряхивают для «перемешивания» двух жидкостей. Раствору дают отстояться и жидкостные фазы сепарируют. После разделения измеряют концентрацию в водной и липидной фазах. Полученное соотношение концентраций называется «коэффициент распределения», что часто не вполне правильно понимают как «жирорастворимость».
2. Важно отметить, что коэффициент распределения, определенный таким образом, будет существенно варьировать в зависимости от:
- pH водной фазы, поскольку она будет определять, какая часть местного анестетика заряжена (более гидрофобна) или не заряжена (более гидрофильна). Значение pH 7,4 типично, и полученный в результате коэффициент распределения называется коэффициент разделения. Коэффициент распределения обычно измеряют, используя местный анестетик, основание и водную фазу pH со значительно более высокой рХа, чем у препарата, что обеспечивает отсутствие заряда в большей части местного анестетика;
- используемого липида. Различные липиды будут давать очень разные коэффициенты распределения и значения, полученные в системе одного растворителя нельзя сравнивать со значениями другой системы. Обращение к «жирорастворимости» препарата без уточнения системы, в которой она определена, будет информацией неполной;
- формы местного анестетика (основание или соль).
В. Связывание с белками. Связывание с белками плазмы варьирует от 5% до 95%. В целом более гидрофобные препараты обладают более высоким связыванием с белками. Фактически свойства препарата, иногда относимые на счет степени «связывания с белками», по-видимому, связаны с его гидрофобностью. Есть ли какие-либо взаимоотношения между связыванием с белками плазмы и белками тканей — неизвестно, и их можно не принимать во внимание.
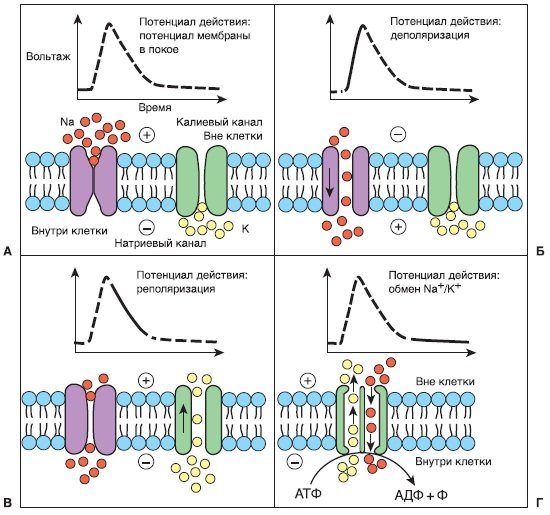
Функция натриевых и калиевых каналов и перемещения иона во время деполяризации нерва. А: В покое натриевый канал закрыт: во внеклеточном пространстве создается относительный избыток ионов натрия (коричневые кружки) и относительный избыток ионов калия во внутриклеточном пространстве (светлые кружки). Поскольку на каждые три положительно заряженных иона натрия во внеклеточном пространстве приходится приблизительно два заряженных иона калия во внутриклеточном пространстве, последнее отрицательно заряжено (от —50 до —90 мВ) относительно внеклеточного. Б: После достаточной стимуляции потенциал-зависимая конфигурация натриевых каналов меняется на открытую и ионы натрия устремляются по своему электрохимическому градиенту внутрь нейрона, что приводит к деполяризации. В: На пике потенциала действия конфигурация натриевого канала спонтанно меняется на неактивное состояние, которое препятствует дальнейшему поступлению натрия и рефрактерно к попыткам повторного открытия в ответ на стимул. Одновременно ворота калиевых каналов открываются и калий устремляется по своему электрохимическому градиенту, компенсируя отрицательный заряд внутри относительно внеклеточной среды (реполяризация). Г: Калий-натриевый насос (Ма+/К+-АТФаза) обменивает три внутриклеточные молекулы натрия на каждые две внеклеточные молекулы калия, таким образом восстанавливая потенциал покоя мембраны и продвигая натриевый канал к закрытой конфигурации. АДФ, аденозиндифосфат; АТФ, аденозинтрифосфат; Ф, фосфат. (Адаптировано из: Barash К, Clitten S. Clinical anesthesia, 3rd edition)
- а1-кислый гликопротеин и альбумин — основные белки плазмы, с которыми связываются местные анестетики. Связывание с этими белками зависит от pH и снижается при ацидозе, поскольку количество способных к связыванию точек в кислой среде снижается.
- В плазме есть несвязанные, или «свободные», фракции местного анестетика, которые способны выходить из плазмы и попадать в другие органы, такие как мозг или сердце. Именно эти свободные фракции ответственны за системную токсичность.
Пациенты с низкой концентрацией белков (например, при нутритивной недостаточности, циррозе, нефротическом синдроме) имеют более высокий риск системной токсичности, чем пациенты с нормальной концентрацией белка в плазме и пациенты с высокой концентрацией белка в плазме (например, некоторые формы рака), которые обеспечивают себе дополнительную степень защиты.
Натриевый канал и проводимость по нерву
- Структура натриевого канала. Натриевый канал млекопитающих — трансмембранный белок, состоящий из трех субъединиц, формирующих чувствительный к потенциалу, натрий-селективный канал через нейрональную мембрану. К настоящему времени идентифицировано десять отдельных генов человека, в которых закодированы десять структурно отличных натриевых каналов. Различные формы представлены в разных тканях (например, мышцах, сердце, центральной и периферической нервной системе). Возможно, существуют мутации, которые повышают или понижают чувствительность к местным анестетикам (такого рода индуцируемые мутации были получены в экспериментальных системах, но в настоящее время клинических подтверждений этому не найдено.
- Проводимость. В покое нейроны поддерживают электрохимический градиент вокруг своих мембран, поскольку Ка+/К+-АТ Фаза (аденозинтри- фосфатаза) закачивает три иона Na+ из аксоплазмы на каждые два иона К+, закачанные в нее. Это значит, что внутренняя среда аксона относительно отрицательно заряжена (от —50 до —90 мВ) и натрия в ней меньше по сравнению с внешней средой. При достаточной «стимуляции» нерва натриевые каналы в очень локальной зоне нервной мембраны открываются, таким образом позволяя ионам Na+ перемещаться в соответствии с их градиентом внутрь аксона и локально «деполяризовать» аксональную мембрану. Если размах деполяризации превышает «порог» (т. е. трансмембранный потенциал снизился достаточно), индуцируется открытие натриевых каналов в прилежащей мембране (это и означает «чувствительность к потенциалу»), что, в свою очередь, индуцирует открытие натриевых каналов, расположенных еще дальше. Таким образом деполяризация распространяется вниз по аксональной мембране, формируя потенциал действия.
- Реполяризация. Через несколько миллисекунд натриевый канал инактивируется зависимым от времени изменением конформации, которое «закрывает» ворота. В инактивированном состоянии натриевый канал не может проводить Na+ и не может открыться вновь при стимуляции (аналогично рефрактерному периоду в миокарде). Сначала потенциал покоя мембраны восстанавливается до нормального за счет внеклеточного перемещения К+, а позднее за счет обмена натрия на калий Na+/K+ АТ Фазой. По мере восстановления потенциала покоя мембраны натриевый канал подвергается дополнительным пространственным изменениям для перехода в закрытое состояние (покоя), во время которого он не проводит ионы Na+, но достаточный стимул (например, деполяризация, сенсорная трансдукция, связывание нейротрансмиттера) будет переводить канал в открытое состояние. Важно заметить, что аффинность местных анестетиков варьирует с изменением состояния натриевых каналов, с максимальным значением в инактивированном состоянии и минимальным в состоянии покоя (закрытом). Эти, зависящие от состояния, различия аффинности лежат в основе «фазного» или «частотно-зависимого» блока. Кроме того, различия между местными анестетиками в степени, с которой они проявляют зависящие от состояния различия аффинности к связыванию, лежат в основе их относительной кардиотоксичности.
- Связывание местных анестетиков. «Рецепторов» для местных анестетиков не существует; правильнее такие участки называть «зонами связывания». Направленные исследования мутагенеза показывают, что зоны связывания местных анестетиков расположены внутри натриевых каналов, рядом с местом их входа в клетку. Местные анестетики блокируют потенциалы действия, либо препятствуя перемещению Na+ по натриевым каналам физически, либо не давая совершить изменения в натриевом канале, необходимые Na+ для того чтобы он мог пройти через пору.
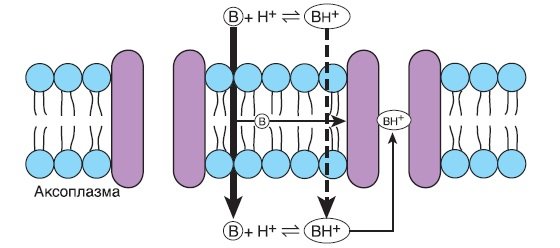
Модель взаимодействия местного анестетика с натриевым каналом. Во внеклеточной жидкости молекула местного анестетика присутствует в равновесном состоянии между нейтральным третичным амином-основанием (В) и положительно заряженным четвертичным амином (ВН+). Незаряженная третичная форма местного анестетика проникает через клеточную мембрану значительно легче, чем заряженная четвертичная форма, но последняя до некоторой степени проникает тоже. Такое же равновесие между незаряженным третичным амином и заряженным четвертичным амином существует и внутри нерва, хотя более низкое значение pH внутри нейрона будет способствовать образованию четвертичной формы в большей степени, чем во внеклеточной жидкости. Только заряженная четвертичная форма способствует взаимодействию точки связывания с местным анестетиком внутри натриевого канала и может реагировать с точкой связывания только изнутри нейрона. Считается, что незаряженные местные анестетики (например, бензокаин) взаимодействуют с натриевыми каналами в разных точках, что возможно только изнутри аксональной мембраны. Как альтернатива, незаряженные местные анестетики могут изменять функцию натриевого канала, изменяя свойства аксональной мембраны и, таким образом, взаимодействие натриевого канала с мембраной.
1. Зона связывания местного анестетика состоит из гидрофобной области, которая принимает гидрофобную часть молекулы местного анестетика для взаимодействия, и гидрофильной области, с которой взаимодействуют четвертичные амины. Аминокислотные замены в этих зонах блокируют эффект местных анестетиков.
Д. Модель действия местного анестетика. В экспериментах in vitro с использованием гигантского аксона кальмара было продемонстрировано, что перманентно заряженные четвертичные амины местных анестетиков обладают относительно слабой местноанестезирующей активностью при нанесении их за пределами мембраны нерва, но вполне сильной при введении в цитоплазму нерва. Напротив, незаряженные третичные амины, введенные внутрь нерва, не очень эффективны как местные анестетики. Эти наблюдения стали основанием следующей модели местноанестезирующего действия третичных аминов.
- Местные анестетики должны проникать через аксональную плазменную мембрану для достижения своих точек связывания.
- Незаряженные, более гидрофобные, третичные амины формируют местные анестетики, легче проникающие через аксональную мембрану.
- Заряженные четвертичные амины формируют местные анестетики, отвечающие за блокаду натриевых каналов.
- Исключения. У этой модели есть несколько исключений.
а. Бензокаин, который не имеет аминогруппы и, следовательно, перманентно не заряжен, тем не менее блокирует натриевые каналы. Бензокаин может иметь различные точки связывания и может попадать на них прямо из плазменной мембраны — вместо аксоплазмы.
б. Перманентно заряженные местные анестетики четвертичные амины (например, тоникаин) in vitro продуцируют медленно развивающуюся, но длительно действующую блокаду натриевых каналов.
Фармакодинамические характеристики in vitro
А. Мощность. Мощность местного анестетика определяется как минимальная локальная концентрация местного анестетика, необходимая для блокады нерва. In vitro на изолированном нерве мощность очень хорошо коррелирует с гидрофобностью. In vivo корреляция все еще есть, но она менее выражена. Кроме того, минимальные блокирующие концентрации in vitro расположены в порядке увеличения или более низки, чем нужные in vivo в связи с поглощением препарата в неспецифических зонах связывания при прохождении через диффузионные тканевые барьеры и подобные препятствия, встречающиеся in vivo.
Б. Частотнозависимый (фазный) блок. Чем быстрее нерв стимулируется in vitro, тем ниже концентрация местного анестетика, необходимая для его блокады. Этот феномен называют частотнозависимым или фазным блоком. Он возникает по разным причинам.
1. Местные анестетики могут достигать своих точек связывания только при открытых каналах. Следовательно, нерв в покое нельзя блокировать, и чем чаще будет происходить стимуляция нерва, тем больше каналов будет открыто для поступления местного анестетика.
2. Аффинность местного анестетика к его точкам связывания наиболее высока в инактивированном состоянии и наименее — в состоянии покоя. Пока канал переходит из инактивированного в состояние покоя, местный анестетик может пройти мимо точки связывания так, что последующая деполяризация находит канал не заблокированным. По мере возрастания запальной частоты каналы все меньше времени находятся в состоянии покоя и, таким образом, остается меньше времени для прохождения местного анестетика мимо точек связывания. На самом деле блокада натриевых каналов происходит в результате баланса между связыванием местного анестетика в инактивированном состоянии и диссоциацией местного анестетика в состоянии покоя.
Фазный блок наблюдается в большей степени у более мощных (гидрофобных) местных анестетиков, поскольку разброс в различиях их аффинности к связыванию между открытым/инактивированным состоянием и состоянием покоя больше, чем у более гидрофильных препаратов. Неясно, до какой степени частотнозависимый блок, легко демонстрируемый in vitro, имеет место in vivo. Тем не менее частотнозависимый блок натриевых каналов миокарда in vivo служит важной причиной того, что гидрофобные местные анестетики более кардиотоксичны, чем гидрофильные.
В. Длина нерва, подвергнутого воздействию, и местноанестезирующий блок. In vitro чем длиннее участок нерва, на который воздействовал местный анестетик, тем ниже концентрация местного анестетика, необходимая для того, чтобы вызвать блокаду. Этот эффект достигает пика при экспозиции на 2,5—3 мм; при длине свыше 3 мм минимальная блокирующая концентрация более не растет.
1. Миелинизированные аксоны. Миелин состоит из плазматических мембран шванновских клеток, обернутых вокруг аксона. На аксоне есть перехваты, которые называют узлами Ранвье, расположенные с фиксированными интервалами между миелинизированными зонами. Миелинизация приводит к значительно более быстрой скорости проведения, поскольку аксональным мембранам нужна только деполяризация в узле. На самом деле деполяризация «перепрыгивает» с узла на узел в процессе, который называется сальтаторное проведение.
а. Местные анестетики могут получать доступ к аксональной мембране миелинизированных аксонов только в узлах Ранвье. In vitro местному анестетику нужно блокировать натриевые каналы приблизительно в трех последовательно расположенных узлах (0,4—4 мм).
а Аксоны человека классифицируют по размеру, функции и наличию или отсутствию миелина. In vitro малые немиелинизированные аксоны наиболее устойчивы к блокаде местными анестетиками, тогда как большие миелинизированные аксоны наиболее чувствительны. In vivo, однако, чувствительность к блокаде местными анестетиками отличается, но причины этого не вполне понятны. «+» означает относительную чувствительность к блокаде местными анестетиками для прекращения аксонального проведения. Значительная вариабельность длин участков связана с тем, что у аксонов большего диаметра длиннее «межузловое» расстояние, чем аксонов с меньшим диаметром. Нужно ли такое же количество узлов для блокады in vivo — неизвестно.
2. Немиелинизированные аксоны. Как и у миелинизированных аксонов, концентрация местного анестетика, необходимая для блокады проводимости по немиелинизированным аксонам, снижается при увеличении длины нерва, подвергнувшегося воздействию местного анестетика.
Г. Тип, размер аксона и местноанестезирующая блокада. Аксоны человека классифицируют в соответствии с их структурой (миелинизирован- ный, немиелинизированный), размером (т. е. диаметром) и функцией (табл. 1.2). Характеристики местноанестезирующей блокады для разных типов аксонов варьируют, но роли размера, миелинизации или функции в аксональной блокаде не вполне ясны.
1. В равновесных условиях in vitro немиелинизированные аксоны (С-волокна) наиболее устойчивы к блокаде местными анестетиками, за ними идут крупные (Аа, Ар) и мелкие (В) миелинизированные аксоны. Легче всего блокировать in vitro миелинизированные аксоны промежуточного размера (AS, Ay). Механизм осуществления таких различий чувствительности точно неизвестен, но ясно, что он связан с размером нерва или миелинизацией per se.
Заключение
Химические особенности и молекулярная фармакология местных анестетиков лежат в основе клинической фармакологии, описанной в последующих главах. Знакомство с принципами, изложенными здесь, облегчит понимание клинической фармакологии различных местных анестетиков при использовании их для специфических блокад. Однако следует помнить, что клиническое действие включает ряд факторов (например, захват, распределение и метаболизм), которых нет в простых системах, используемых для исследования химии и фармакологии на клеточном уровне. В связи с этим последующие главы крайне важны для понимания клинического применения этого важного класса препаратов.